
摘要:[目的]子代窝群大小和性比是聚寄生蜂在寄主上产卵的重要决策,寄主资源量是影响该决策的主要因素之一。本研究旨在探明准社会性硬皮肿腿蜂属(Sclerodermus)的生殖策略。[方法]以管氏肿腿蜂(S. guani Xiao et Wu)为对象,以不同体质量的青杨天牛(Saperda populnca L.)幼虫为寄主,人为控制窝群大小(初孵幼虫数量),设置不同水平处理,观测子代蜂的存活率、性比和体型大小,以推测最适的生殖策略。[结果]在自然窝群(大于等于50头)下,子代蜂存活至成虫期的比例随寄主体质量增大而提高;但在人为减小窝群的几个处理(10、20、30和40头)中,子代蜂发育至成虫羽化的存活率均很高,且与寄主体质量无关,4个处理之间亦没有显着差异。窝群大小不影响子代蜂性比,寄主体质量也无影响,性比均始终保持极端偏雌的比例。羽化出的成虫雌蜂体质量随子代蜂窝群增大而线性减小,而且该趋势不受寄主体质量的影响。[结论]管氏肿腿蜂可能采用“产固定性比且略大的窝群”的生殖策略。
关键词:准社会行为; 聚寄生蜂; 体外寄生蜂; 窝卵决策; 性比;
聚寄生蜂在产卵时作出的主要决策之一是,在1头寄主上产多少粒卵———窝卵数(clutch size)。根据Lack最适窝卵数假说预测,寄生蜂所产窝卵数应使其从中获得的适合度最大化[1]。对许多种类寄生蜂的研究发现,雌蜂(母蜂)所产窝卵数与寄主体型大小呈正相关线性关系,支持最适窝卵数假说[2]。但从子代适合度视角看,由于寄主资源量是固定的,故子代蜂所获得的适合度随窝卵数增大而减小。所以,产卵雌蜂与子代蜂之间在适合度利益上存在矛盾,即所谓“亲子冲突(parent-offspring conflict)”,但在自然选择压力下演化出的进化稳定策略使双方的利益达到某种动态平衡[3,4,5]。对于聚寄生蜂而言,除了窝卵数外,子代性比是另一重要决策,因为雌蜂可控制子代蜂的性别:产二倍体的受精卵为雌性,产单倍体的卵为雄性。由于聚寄生蜂营近亲繁殖,根据“Hamilton局部配偶竞争”理论预测,在无同类雌蜂竞争的情况下产偏雌性比的子代蜂群;在有同类竞争的情况下,增大雄性比例[6]。在硬皮肿腿蜂属(Sclerodermus)中,雌蜂始终产极端偏雌性比的子代蜂蜂群[7]。迄今,对这类营准社会行为的寄生蜂的生殖策略,知之甚少。
硬皮肿腿蜂(肿腿蜂科Bethylidae)是树木蛀干害虫的重要寄生性天敌,在我国广泛用于森林蛀干害虫天牛的生物防治[8,9,10]。自从1928年Wheeler[11]首次记载硬皮肿腿蜂具有抚育子代行为以来,对硬皮肿腿蜂的研究主要围绕生物防治应用进行。近年来,对其抚育行为的理论研究日渐增多,发现母蜂(交配过的雌蜂)抚育对子代蜂顺利完成发育具有重要作用[12,13],但母蜂因此可能失去寻找新寄主的机会[14]。Tang等[7]针对母蜂之间合作抚育的机制提出了一个新的假说———母蜂合作有利于制服并寄生利用体型较大的寄主,并通过试验初步证实了该假说。若干接蜂试验研究结果发现,在管氏肿腿蜂(Sclerodermus guani Xiao et Wu)中合作利用寄主的母蜂所分摊的子代蜂数少于其单独寄生所得到的子代蜂数[7,14]。导致这一差异的原因是多方面的,例如:合作的母蜂控制产卵数量;合作的母蜂在抚育中控制子代存活数(如:杀幼);子代蜂在发育中相互竞争;或者同时采用以上几个生殖策略。在当前尚无法鉴别母蜂与子代蜂亲缘关系的情况下,难以明确鉴别肿腿蜂采用哪一生殖策略。目前可行的方法是通过人为控制子代蜂数量,观测子代蜂发育表现,从而推测雌蜂单独寄生时的生殖策略:如果母蜂仅根据寄主大小来决定产卵量,则子代蜂在不同体型大小的寄主上都具有良好的发育表现,这一策略为某些单独寄生、不抚育子代的聚寄生蜂采用[2];如果母蜂不能精准控制产卵量,而是采用通过子代蜂竞争来控制蜂群数量的生殖策略,则子代蜂发育表现完全依赖于寄主体型大小(即营养资源多少),这一策略为单独抚育子代的棱角肿腿蜂[Goniozus nephantidis(Muesebeck)]所采用[15]。在可合作抚育子代的硬皮肿腿蜂中,根据非控制性(不控制窝蜂数)试验观察结果推测,可能采取后一生殖策略[7,16,17],但尚需控制性试验予以验证。
为探究硬皮肿腿蜂雌蜂的生殖策略,本研究以管氏肿腿蜂及其寄主青杨天牛(Saperda populnca L.)为对象,在不同体质量的寄主上,通过人为减少子代幼虫数量设置窝群大小处理,观察羽化出蜂数、性比和雌蜂体型大小。
1、材料与方法
1.1、供试昆虫
供试寄主青杨天牛(Saperda populnca L.)由江苏省林业科学院森林保护研究所提供,均为从杨树枝中剖出的越冬老熟幼虫。供试管氏肿腿蜂(Sclerodermus guani Xiao et Wu)来自江苏省林业科学院森林保护研究所林业有害生物天敌繁育室,近年来用于江苏等省部分林区防治松墨天牛。试虫均在4℃恒温下冷藏保存备用。
1.2、试验方法
称量青杨天牛幼虫(Mittler XS105 Dual Range,精确到10×10-4g)后,单头接入指形管(直径1 cm、长度4 cm)中,再释放1头交配过的管氏肿腿蜂雌蜂(以下简称“母蜂”)。当所产卵孵化后,通过移除初孵幼虫设置以下子代蜂数量梯度:10、20、30、40头,以自然产卵、孵化的幼虫数量(大于等于50头)为对照。每天观察记录幼虫数直至其化蛹结茧,记录幼虫发育历期(从首次观察到幼虫到第1个完整茧出现的时间);子代蜂羽化后,计数总出蜂量,并将所有子代蜂雌成虫集体称量以获得个体平均体质量,作为子代蜂体型大小的测度。试验在养虫室内进行,温度(26±1)℃。每个处理重复20次。
1.3、数据分析
用logistic广义线性模型(二项分布,连接函数logit)分析不同数量子代蜂幼虫处理下发育至成虫羽化的存活概率和性比(雄虫占子代蜂总数的比例)随寄主体质量增大的变化趋势,当数据的方差大于模型假定的方差(过度离散)时,用拟二项分布(quasi-binomial distribution)予以校正。处理因素(幼虫数量和寄主体质量)的影响显着性用对数似然比卡方测验进行检验。用一般线性回归分析子代蜂体质量,处理因素的影响显着性用F测验进行检验。数据分析用R软件[18]。
2、结果与分析
2.1、子代蜂窝群大小和寄主体质量对子代蜂存活率的影响
子代蜂窝群大小显着影响子代蜂存活率(χ2=79.74,P<0.001),寄主体质量对子代蜂存活率无显着影响(χ2=3.20,P=0.07),窝群大小与寄主体质量之间存在显着互作影响(χ2=9.78,P=0.04)。在窝群为10、20、30和40头处理中,子代蜂存活率均很高,而且均不受寄主体质量的影响(10头:z=0.02,P=0.89;20头:z=1.54,P=0.22;30头:z=0.02,P=0.89;40头:z=0.25,P=0.62);在自然窝群(大于等于50头)下,子代蜂存活率总体低于以上处理(z=11.159,P<0.001),寄主体质量每增加1 mg子代蜂存活率增加4%(图1)。
2.2、子代蜂窝群大小和寄主体质量对子代蜂性比的影响
子代蜂窝群大小(χ2=1.955,P=0.744)、寄主体质量(χ2=0.418,P=0.518)及其两因素互作(χ2=6.212,P=0.184)均对性比(雄性占比)无显着影响,性比平均为5%(图2)。
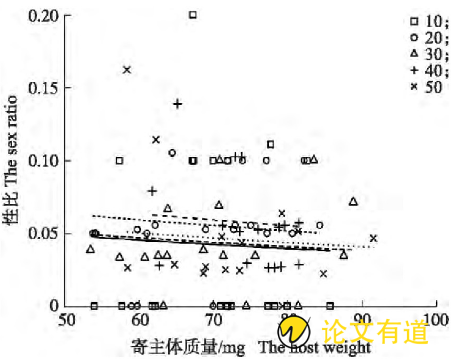
图2 不同窝群大小的子代蜂性比与寄主体质量的关系
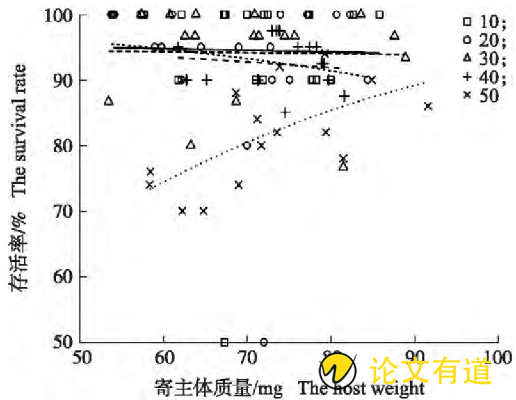
图1 不同窝群大小处理下子代蜂存活率与寄主体质量的关系
2.3、子代蜂窝群大小和寄主体质量对子代雌蜂体型大小的影响
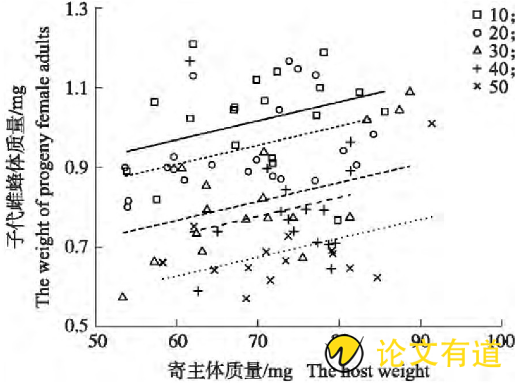
图3 不同窝群大小处理下的子代雌蜂体质量与寄主体质量的关系
子代蜂窝群大小(F4,83=24.69,P<0.001)与寄主体质量(F1,83=11.69,P<0.001)均显着影响子代雌蜂体质量,两因素间无显着互作影响(F4,79=1.89,P=0.12)(图3)。在子代窝群大小不变的情况下,子代雌蜂体质量随寄主体质量增加而直线增加,寄主体质量每增加1 mg,子代雌蜂体质量增加0.005 mg(t=3.42,P<0.001);在寄主体质量不变的情况下,子代雌蜂体质量随子代窝群增大而降低,与窝群为10头的子代雌蜂体质量相比,窝群为20头的无显着差异(P<0.01),其他窝群的雌蜂体质量分别减小0.202 mg(30头,t=5.33,P<0.001)、0.240 mg(40头,t=6.14,P<0.001)和0.343 mg(大于等于50头,t=8.67,P<0.001);与窝群为20头的处理相比,其他窝群的子代雌蜂体质量分别减小0.141 mg(30头,t=3.75,P=0.003)、0.179 mg(40头,t=4.57,P<0.001)和0.282 mg(大于等于50头,t=7.15,P<0.001);与窝群为30头蜂处理相比,大于等于50头窝群下的子代雌蜂体质量减小0.140 mg(t=3.42,P=0.01)。
3、讨论
本研究发现,在人为减少管氏肿腿蜂所产子代幼虫数量(小于50头)的处理中,子代蜂发育至成虫羽化的存活率均很高,而且不受寄主体质量的影响;但在自然窝群(大于等于50头)下,子代蜂存活率总体较低,随寄主体质量增大而增大。根据该结果推测,管氏肿腿蜂雌蜂可能采用“适量超额产卵”对策,“适量”是指产卵量略多于存活至羽化出蜂的量。例如,在每头松墨天牛(Monochamus altematus Hops)幼虫上接2、3或4头管氏肿腿蜂情况下,每头母蜂平均产49、44和24粒卵,存活至成虫羽化的比例分别为63.3%、81.8%和91.7%[19]。本研究发现,子代雌成虫体型大小随子代虫量增多而线性减小,这说明子代蜂对寄主资源存在明显的竞争。该反比关系广泛存在于无抚育习性的聚寄生蜂中[20],也存在于肿腿蜂的近缘属棱角肿腿蜂中[15]。
采用“适量超额产卵”的对策,有以下适应性:1)母蜂通过多产子代可提高与其他雌蜂的相对竞争力。雌蜂通过合作可有效利用体型较大的寄主[13],但在制服寄主后,子代蜂面临着竞争,故适当多产卵可提高与其他母蜂的子代进行竞争的优势。2)即使雌蜂单独寄生寄主,采用该生殖策略可通过子代的竞争达到择优劣汰的效果。3)在一个寄主资源量不确定性很大的环境中,一旦找到寄主适当多产卵可能降低来自不确定性的风险。肿腿蜂抱卵量远超过其产卵量,但由于抚育子代、寻找寄主需耗费大量时间,可能难以实现其生殖潜力[12],在这种情况下,适当多产卵比浪费卵更有利。当然,母蜂抚育可提高子代的总体发育表现[10,11],尤其是当寄主质量严重恶化的情况下,母蜂可取食子代卵以避免过渡消耗寄主[21]。
本研究发现,子代幼虫量和寄主体型大小均不影响子代出蜂性比。肿腿蜂子代蜂性比通常比较稳定[7,10,14,20]。采用极端偏雌的性比分配策略,可能有助于增强子代雌蜂搜寻和利用寄主[13],符合资源强化假说的预测[22]。
综上所述,管氏肿腿蜂雌蜂可能采用“产固定性比且略大的窝群”的生殖策略。该初步结论有待于采用能精确鉴别肿腿蜂亲子关系的分子生物学技术予以进一步验证。
参考文献
[1] Charnov E L,Skinner S W. Complementary approaches to the understanding of parasitoid oviposition decisions[J]. Environmental Entomology,1985,14:383-391.
[2] Godfray H C J. Parasitoids:Behavior and Evolutionary Ecology[M]. Princeton,New Jersey:Princeton University Press,1994.
[3] Skinner S W. Clutch size as an optimal foraging problem for insects[J]. Behavioral Ecology and Sociobiology,1985,17(3):231-238.
[4] Charnov E L,Skinner S W. Evolution of host selection and clutch size in parasitoid wasps[J]. Florida Entomologist,1984,67:5-21.
[5] Charnov E L,Skinner S W. Clutch size in parasitoids:the egg production rate as a constraint[J]. Evolutionary Ecology,1988,2(2):167-174.
[6] Hamilton W D. Extraordinary sex ratios[J]. Science,1967,156(3774):477-488.
[7] Tang X Y,Meng L,Kapranas A,et al. Mutually beneficial host exploitation and ultra-biased sex ratios in quasisocial parasitoids[J]. Nature Communications,2014,5(12):4942.
[8] 徐克勤,徐福元,王敏敏,等.应用管氏肿腿蜂防治松褐天牛[J].南京林业大学学报(自然科学版),2002,26(3):48-52.Xu K Q,Xu F Y,Wang M M,et al. The techniques of Scleroderma guani Xiao et Wu to control pine sawyer beetles[J]. Journal of Nanjing Forestry University(Natural Sciences Edition),2002,26(3):48-52(in Chinese with English abstract).
[9] 姚万军,杨忠岐.人工繁殖管氏肿腿蜂的替代寄主研究[J].中国生物防治,2008,24(3):220-226.Yao W J,Yang Z Q. Mass-rearing of Sclerodermus guani(Hymenoptera:Bethylidae)with substitute host[J]. Chinese Journal of Biological Control,2008,24(3):220-226(in Chinese with English abstract).
[10] 祁伟.管氏肿腿蜂防治锈斑楔天牛试验初报[J].陕西林业科技,2009(2):89-91.Qi W. A Primary report on the control of Saperda balsamifera Motschulsky by Scleroderma guani[J]. Shaanxi Forest Science and Technology,2009(2):89-91(in Chinese with English abstract).
[11] Wheeler W M. The Social Insects:Their Origin and Evolution[M]. London:K.Paul,Trench,Trubner,1928.
[12] Hu Z J,Zhao X L,Li Y S,et al. Maternal care in the parasitoid Sclerodermus harmandi(Hymenoptera:Bethylidae)[J]. PLoS One,2012,7(12):e51246.
[13] 伍绍龙,徐福元,李保平,等.管氏肿腿蜂雌性抚育中幼虫转移行为的启动和节律[J].昆虫学报,2013,56(4):392-397.Wu S L,Xu F Y,Li B P,et al. Initiation and rhythm of larva translocation behavior during maternal care in an ectoparasitoid Sclerodermus guani(Hymenoptera:Bethylidae)[J]. Acta Entomologica Sinica,2013,56(4):392-397(in Chinese with English abstract).
[14] 伍绍龙,周志成,彭曙光,等.管氏肿腿蜂抚育行为的代价和收益[J].中国生物防治学报,2017,33(1):39-43.Wu S L,Zhou Z C,Peng S G,et al. Costs and benefits of maternal care behavior in the parasitoid Sclerodermus guani(Hymenoptera:Bethylidae)[J].Chinese Journal of Biological Control,2017,33(1):39-43(in Chinese with English abstract).
[15] Hardy I C W,Grifiths N T,Godfray H C J. Clutch size in a parasitoid wasp:a manipulation experiment[J]. The Journal of Animal Ecology,1992,61:121-129.
[16] 林芳芳,唐秀云,孟玲,等.寄主体型大小和母蜂数对管氏肿腿蜂产卵前期和发育历期的影响[J].南京农业大学学报,2015,38(4):584-589. DOI:10.7685/j.issn.1000-2030.2015.04.009.Lin F F,Tang X Y,Meng L,et al. Pre-oviposition and developmental duration in response to host body size and numbers of foundresses in Sclerodermus guani(Hymenoptera:Bethylidae)[J]. Journal of Nanjing Agricultural University,2015,38(4):584-589(in Chinese with English abstract).
[17] 周冰颖,李保平,林芳芳,等.寄主体型大小对管氏肿腿蜂生殖潜力的影响[J].昆虫学报,2016,59(3):316-321.Zhou B Y,Li B P,Lin F F,et al. Influence of host body size on potential reproductive capability of Sclerodermus guani(Hymenoptera:Bethylidae)[J].Acta Entomologica Sinica,2016,59(3):316-321(in Chinese with English abstract).
[18] R Core Team. R:A language and environment for statistical computing[OL]. R Foundation for Statistical Computing,Vienna,Austria. 2016.
[19] 吴伟,程绍传,刘德波.松墨天牛幼虫繁育肿腿蜂适宜蜂虫比研究[J].西南林学院学报,2008,28(3):24-29.Wu W,Cheng S C,Liu D B. Study on the spatial distribution pattern of Blastesthia sp.[J]. Journal of Southwest Forestry College,2008,28(3):24-29(in Chinese with English abstract).
[20] Kapranas A,Hardy I C W,Tangled X,et al. Sex ratios,virginity,and local resource enhancement in a quasisocial parasitoid[J]. Entomologia Experimentalis et Applicata,2016,159:243-251.
[21] 武辉,王小艺,李孟楼,等.白蜡吉丁肿腿蜂的生物学和生态学特性及繁殖技术研究[J].昆虫学报,2008,51(1):46-54.Wu H,Wang X Y,Li M L,et al. Biology and mass rearing of Sclerodermus pupariae Yang et Yao(Hymenoptera:Bethylidae),an important ectoparasitoid of the emerald ash borer,Agrilus planipennis(Coleoptera:Buprestidae)in China[J]. Acta Entomologica Sinica,2008,51(1):46-54(in Chinese with English abstract).
[22] West S A. Sex Allocation[M]. Princeton:Princeton University Press,2009.