
摘要:采用盆栽试验研究比较2个黑麦草品种在不同Cd污染水平 (0、75、150、300、600 mg/kg) 下生长量、抗氧化酶活性、Cd含量及积累量的差异,以筛选出更适合作为土壤镉污染修复的品种。结果表明:随着土壤镉水平的增加,2个黑麦草品种根系干重、‘阿伯德’地上部干重和植株总干重呈先增加后降低的趋势。土壤镉污染水平为75 mg/kg时‘,阿伯德’地上部和植株总干重达到最大值(10.92 g/盆和12.03 g/盆),较对照分别增加了11.13%和10.67%.2个黑麦草品种各部位SOD活性、CAT活性和根系POD活性,均随土壤镉水平的增加呈先升高后降低的趋势,地上部MDA含量则逐渐升高。2个黑麦草品种镉转运系数、镉富集系数均大于1.黑麦草对Cd耐性和吸收富集存在基因型差异。比较2个黑麦草品种,在75~600 mg/kg Cd胁迫下,‘阿伯德’品种地上部干重和总干重均高于‘邦德’品种,且‘阿伯德’地上部和根系镉含量及镉积累量高于‘邦德''.阿伯德’更适合作为土壤镉污染的植物修复材料。
关键词:土壤镉污染; 黑麦草; 镉吸收; 转运系数; 富集系数;

0 引言
随着矿山的不断开发和尾矿的日积月累,以及交通和工业的快速发展、农药化肥的滥用等因素,土壤镉 (Cadmium, Cd) 污染问题日趋严重[1,2,3,4,5].Li等[6]对中国22省市中72个矿区土壤重金属污染状况调查显示Cd平均值比中国II级土壤环境质量标准高36.5倍。韦妮玉等[7]研究了广西典型矿区的矿业活动对农田Cd污染的累积影响,通过采集12个自然屯农田156土壤样品分析发现,156个土壤样品中镉超标率为98.1%.重金属Cd污染对于农业生产和生态环境保护均会产生不良影响,重金属污染修护成为了不可忽视的问题。植物修复重金属污染土壤作为一种新兴的环境治理技术,是土壤重金属Cd污染治理和修复的主要方法之一[8].植物修复技术适用于大范围Cd污染土壤,修复潜力巨大且可以美化环境,在实际修复和治理Cd污染土壤中具有较强的现实意义。
黑麦草 (Lolium spp.) ,属禾本科 (Poaceae) 黑麦草属,具有生长快,分蘖再生能力强,产量高,耐寒、耐旱等特点,此外,黑麦草能富集重金属,在重金属污染土壤上种植有很强的抗性,还能在重金属污染较严重、环境恶劣的尾矿地区生存,甚至正常生长[9,10].李希铭[11]分析36种常见草本植物和2种超富集植物在Cd胁迫下Cd耐性和富集特性差异发现,草本植物对低浓度Cd有较高的耐性,一年生黑麦草 (Lolium multiflorum L.) 地上部Cd积累量最高,分别是超富集植物龙葵、印度芥菜的3.4倍和4.1倍。
同种植物不同品种之间对Cd的吸收和积累存在较大差异[12].目前有关不同品种黑麦草对Cd胁迫的抗性,Cd的吸收和积累差异研究较少[13].因此,本试验以2个黑麦草品种(‘邦德’和‘阿伯德’)为材料,通过土培试验,研究2个黑麦草品种在不同Cd污染水平 (0、75、150、300、600 mg/kg) 土壤上生长、抗氧化酶活性、Cd含量及积累量的差异,以期为土壤镉污染植物修复提供资料积累。
1 材料与方法
1.1 供试材料
供试土壤采自重庆市北碚区西南大学紫色土基地。土壤pH 5.29,有机质14.70 g/kg,全氮0.65 g/kg,碱解氮46.83 mg/kg,有效磷33.56 mg/kg,速效钾69.30 mg/kg,全镉0.11 mg/kg,未检测到有效镉。黑麦草 (Lolium multiflorum L.) ,通过前期试验筛选出2个生长和镉含量差异明显的品种(‘邦德’和‘阿伯德’)进行土培试验。邦德种子购买于江西稻草人农业园,阿伯德种子购买于郑州华丰草业科技。
1.2 试验设计
土培试验时间为2016年3月7日-2016年5月29日,地点在西南大学资源环境学院1号温室内。试验设置5个Cd水平:0、75、150、300、600 mg/kg(前期试验确定),外源镉以溶液形式加入土壤,Cd源为CdCl2·2.5H2O.具体操作方法为称取5 kg过5 mm筛风干土壤于避光塑料盆中,人工向土壤添加CdCl2·2.5H2O溶液,充分混匀。采用自制根袋法[14],根袋(高×周长=12 cm×20 cm)装过2 mm筛的风干土200 g作为根际土,4.80 kg土装入黑色塑料盆中(直径25 cm,高17 cm)并置于根袋外围作为非根际土。平衡2周后,移栽生长一致的植株(高15 cm)幼苗于根袋中,‘邦德’和‘阿伯德’各40株/盆。底肥为N 180 mg/kg, P2O5 100 mg/kg, K2O150 mg/kg,肥源及具体操作详见参考文献[1,2].每个处理3次重复,随机排列。黑麦草生长60天收获。
1.3 分析方法
1.3.1植株地上部和根系镉含量测定
黑麦草地上部和根系镉含量测定采用HNO3:HClO4为4:1的混合酸消煮,采用原子吸收分光光度法测定 (Perkin Elmer SIMMA 6000, Norwalk, USA) .检测限为0.005 mg/kg.采用国家标准与技术研究所提供的植株标准物质 (GBW#08513) 对测定结果进行质量监控。所有植物样品的Cd回收率均高于95%,相对标准偏差 (RSD) 的精度在10%以内。
部分指标计算公式如 (1) [4,15]~ (4) [16]所示。
1.3.2抗氧化物质和保护酶活性测定
超氧化物歧化酶 (SOD) 、过氧化物酶 (POD) 采用文献[16]中的方法测定, 过氧化氢酶 (CAT) 活性和丙二醛 (MDA) 含量采用公式[17]中的方法测定。
1.4 数据处理与统计分析
采用Microsoft Excel 2010软件对数据进行处理,采用SPSS 21.0进行方差分析。
2 结果与分析
2.1 镉胁迫对黑麦草生物量的影响
从表1可以看出,2个品种黑麦草地上部和根系干重在不同Cd处理差异达到了显着水平 (P<0.05) .随着土壤镉水平的增加,‘邦德’地上部干重和植株总干重降低,根系干重先增加后降低;‘阿伯德’地上部、根系、植株干重均先增加后降低。在土壤镉污染水平为75 mg/kg时,‘阿伯德’地上部、植株干重显着提高,地上部和总植株干重达到最大值(10.93 g/盆和12.03 g/盆),分别较对照增加了11.1%、10.7%;‘阿伯德’根系干重在150 mg/kg Cd水平下达到最高,较对照增加了43.6%.随土壤镉水平的增加,‘邦德’地上部干重和总干重逐渐降低,在75~600 mg/kg Cd胁迫下,地上部干重和植株总干重分别较对照降低了19.3%、36.1%、75.9%、87.8%和19.8%、29.9%、73.2%、86.9%;而‘邦德’根系在150 mg/kg Cd水平下达到最大值(1.79 g/盆),较对照增加了9.8%.比较2个品种黑麦草,在75~600 mg/kg Cd胁迫下,‘阿伯德’地上部干重和总干重均高于‘邦德’品种;在高镉处理条件下 (300~600 mg/kg Cd) ,‘阿伯德’根系干重高于‘邦德’品种。
2.2 镉胁迫对黑麦草抗氧化酶活性的影响
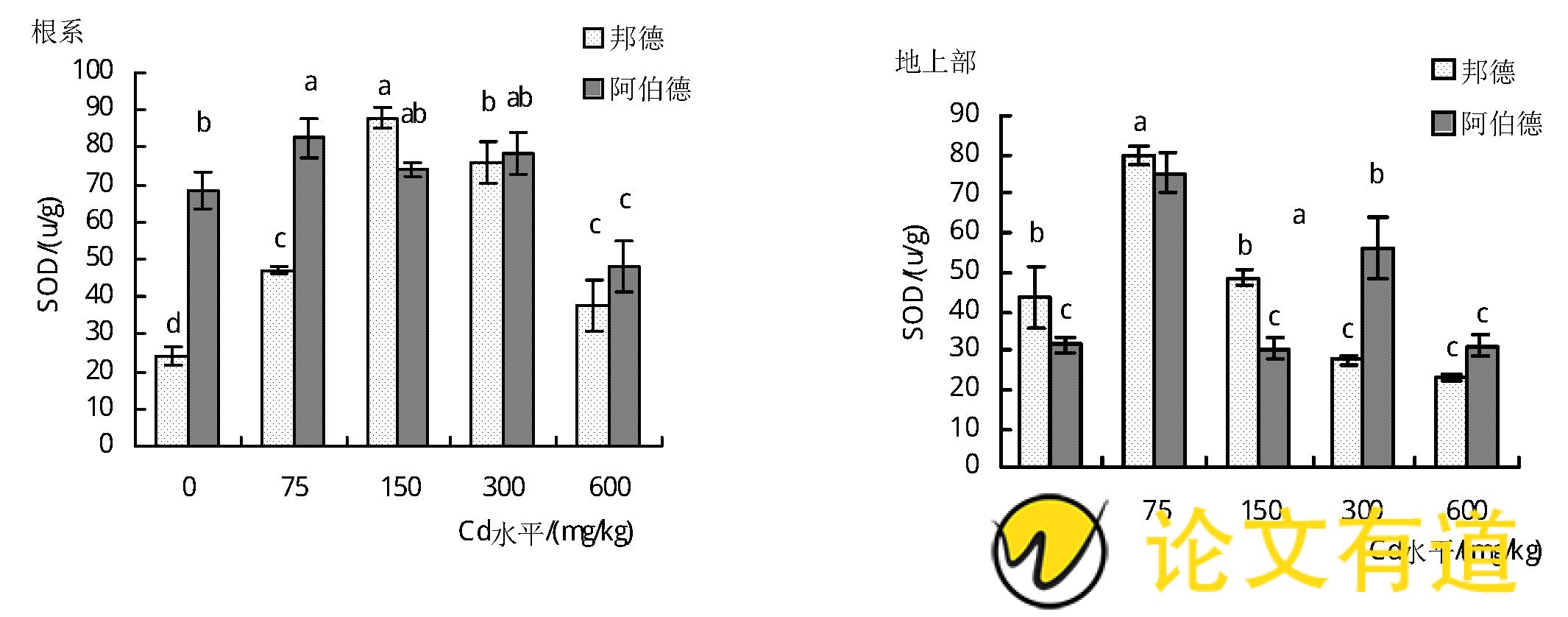
由图1可知,随着土壤镉水平的增加,‘邦德’品种黑麦草根系SOD活性呈显着增加趋势 (P<0.05) ,增幅为7.9%~267.8%.2个品种黑麦草地上部和‘邦德’根系的SOD酶活性均呈先升高后降低的趋势。镉处理水平为75 mg/kg时,‘邦德’和‘阿伯德’地上部SOD活性达到最大值,分别为80.12 u/g和75.51 u/g,较对照增加了83.6%和139.1%.而‘邦德’和‘阿伯德’分别在150 mg/kg和75 mg/kg Cd水平时,根系SOD活性最高,与对照相比分别增加了267.8%和21.8%.
表1 不同镉水平土壤对黑麦草干重的影响
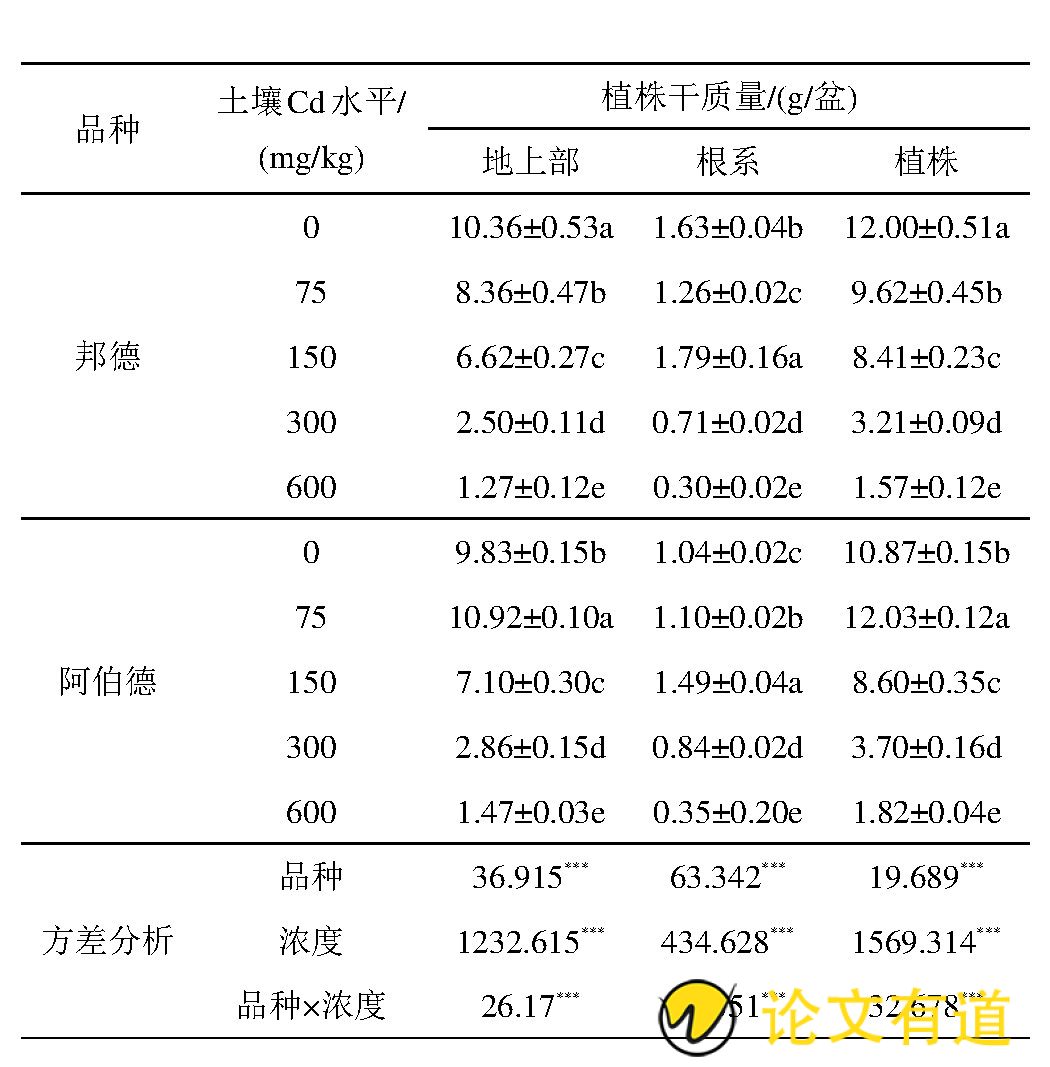
注:所列结果为平均值±标准偏差。表中不同小写字母表示同一品种不同镉处理之间差异达0.05%的显着水平 (P<0.05) .“*”表示0.01<P<0.05、“**”表示0.001<P<0.01、“***”表示P<0.001.下同。
由图2可知,随着土壤镉水平的增加,2个黑麦草品种地上部POD酶活性均呈先降低后升高的趋势 (P<0.05) .当土壤镉水平为300 mg/kg时地上部POD活性达到最大值,‘邦德’和‘阿伯德’地上部POD活性与对照相比分别增加了138.6%和104.7%.而‘邦德’和‘阿伯德’根系POD活性则呈先升高后降低的趋势,在土壤镉水平为75 mg/kg时最大,分别较对照增加了43.6%和22.1%,其后随着土壤镉水平的增加,POD活性显着降低至低于对照水平。
图1 镉胁迫对黑麦草SOD活性的影响
由图3可知,随着土壤镉水平的增加,2个品种黑麦草地上部和根系CAT酶活性均呈先升高后降低的趋势。其中‘邦德’和‘阿伯德’地上部CAT活性在75 mg/kg Cd水平时达到最大值,其中,地上部CAT活性分别为230.5和220 u/ (g·min) ,是对照的1.8倍和1.3倍;根系CAT活性分别为114和141.2 u/ (g·min) ,分别是对照的4倍和3.6倍。
2.3 镉胁迫对黑麦草MDA含量的影响
丙二醛 (MDA) 是衡量逆境胁迫对植物伤害程度的重要生理指标。由图4可知,在镉处理下2个黑麦草品种地上部(除75 mg/kg Cd水平下‘阿伯德'MDA含量)MDA含量均显着高于对照 (P<0.01) ,’邦德‘品种增幅为140.0%~379.3%,在Cd水平为300 mg/kg时达到最高;’阿伯德‘品种在Cd水平为600 mg/kg时达到最高,是对照的3.9倍。随着土壤镉水平的增加,’邦德‘和’阿伯德‘根系MDA含量呈先升高后降低的趋势,当镉处理水平为75 mg/kg时达到最高,分别较对照增加了81.5%和111.1%.
2.4 镉胁迫对黑麦草Cd含量与Cd积累量的影响
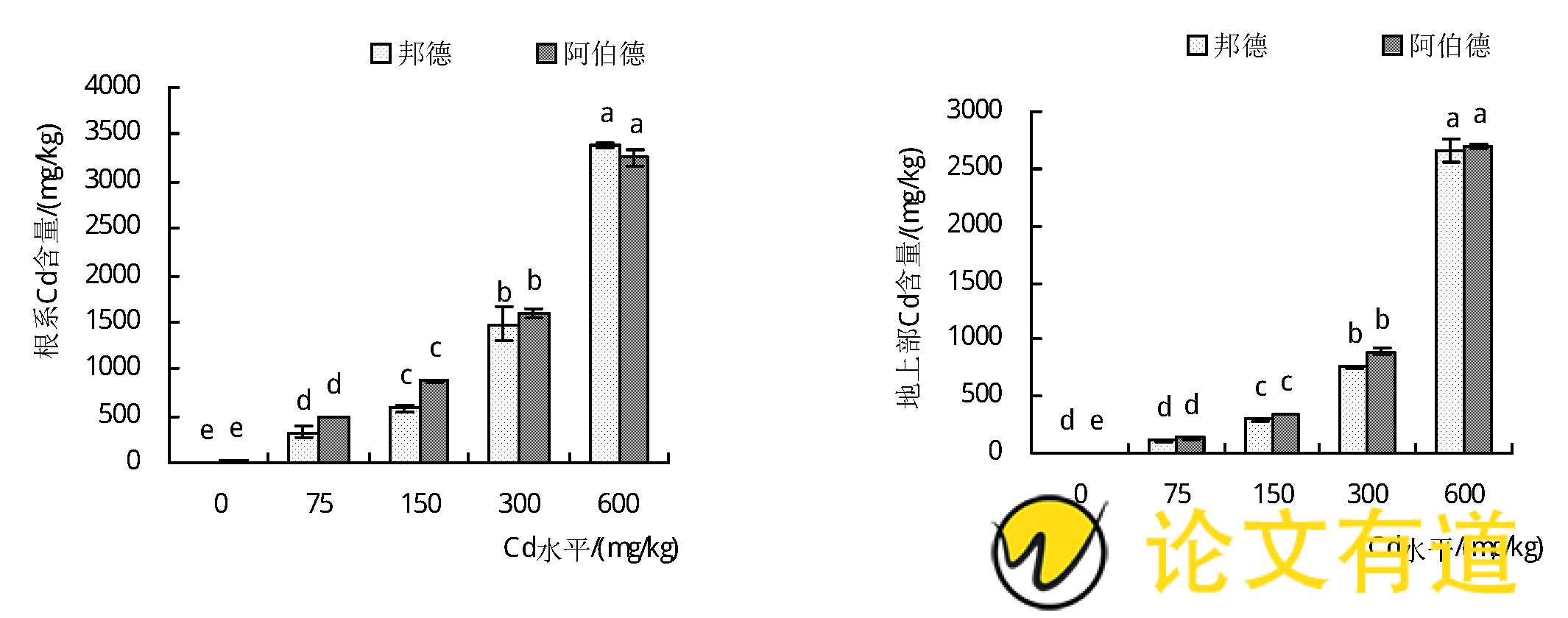
从图5可以看出,2个黑麦草品种地上部和根系镉含量随着镉处理水平的增加呈显着上升趋势 (P<0.05) .在同一镉胁迫条件下,2个黑麦草品种地上部、根系镉含量顺序为根系>地上部。当土壤Cd水平为75 mg/kg时,’邦德‘和’阿伯德‘地上部Cd含量分别为111.19、133.69 mg/kg,均已超过镉超富集植物临界值 (100 mg/kg) .黑麦草对镉富集能力较强,在75~600 mg/kg Cd胁迫下,’邦德‘和’阿伯德‘地上部和根部镉含量分别是对照的30.2~787.5倍和18.0~420.5倍。比较2个黑麦草品种之间地上部和根系镉含量,除600 mg/kg Cd胁迫下’阿伯德‘根系镉含量低于’邦德‘品种,其余处理下’阿伯德‘地上部、根系镉含量均高于’邦德‘,其中’阿伯德‘地上部镉含量在Cd75、Cd150、Cd300和Cd600条件下分别是’邦德‘的1.2、1.1、1.2、1.0倍;在Cd75、Cd150和Cd300条件下,’阿伯德‘根系镉含量分别是’邦德‘的1.5、1.5、1.1倍。说明’阿伯德‘品种黑麦草对镉的富集能力强于’邦德‘品种。
图2 镉胁迫对黑麦草POD活性的影响
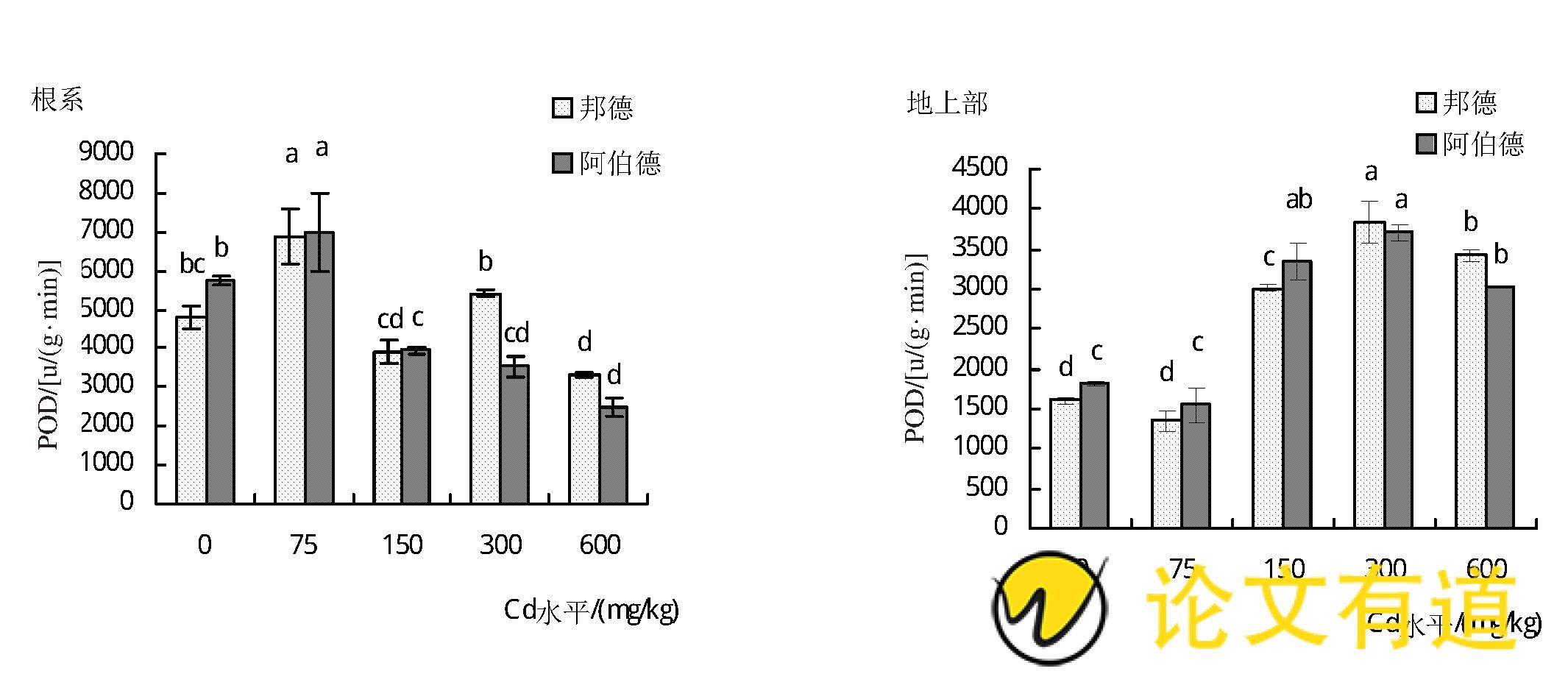
图3 镉胁迫对黑麦草CAT活性的影响
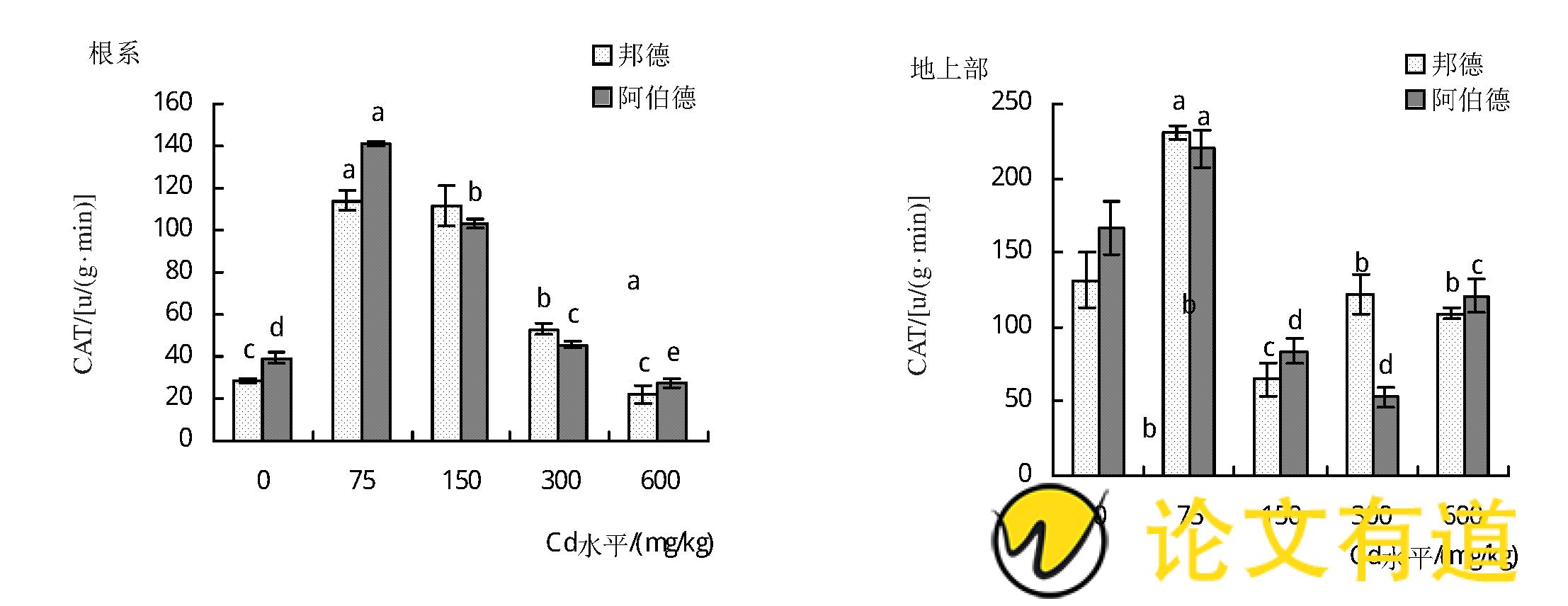
图4 镉胁迫对黑麦草MDA含量的影响
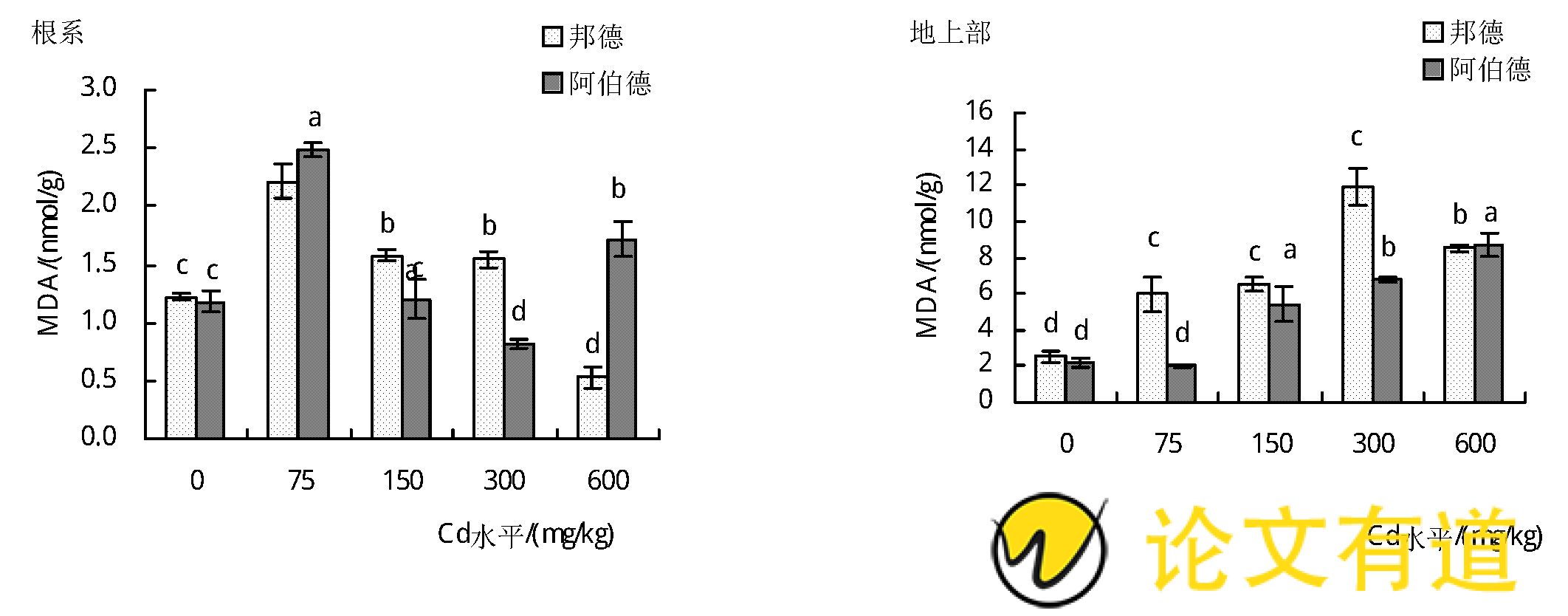
图5 不同Cd污染水平土壤对黑麦草Cd含量的影响
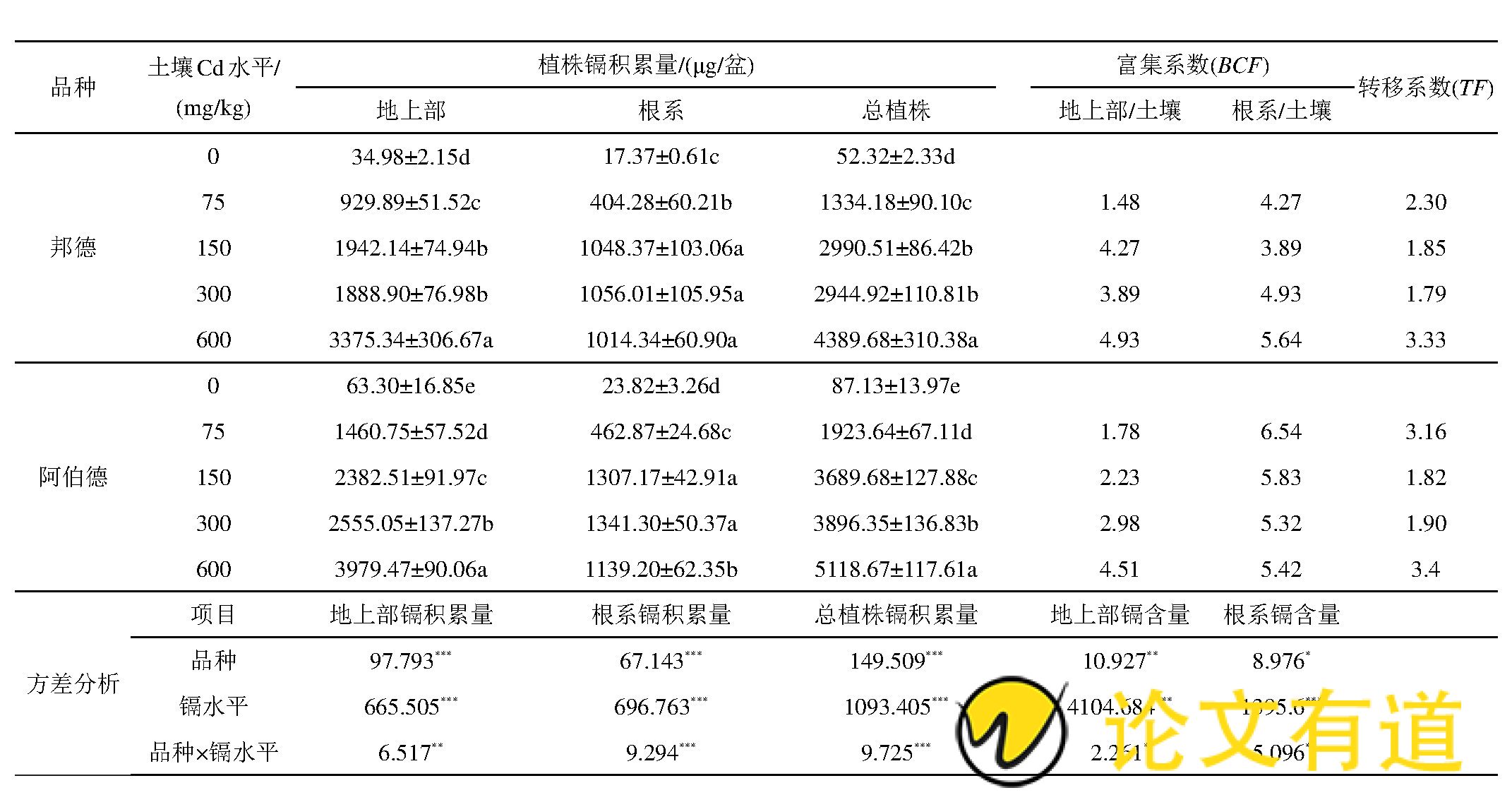
如表2所示,随着镉处理水平的增加,2个黑麦草品种地上部和植株总镉积累量也显着增加 (P<0.05) ,’邦德‘品种地上部、总镉积累量分别是对照的26.6~96.5、25.5~83.9倍;’阿伯德‘品种地上部、总镉积累量分别是对照的23.1~62.9、22.1~58.8倍。’邦德‘和’阿伯德‘根系镉积累量在300 mg/kg Cd胁迫下达到最高,分别是对照的60.8、56.3倍。’邦德‘和’阿伯德‘镉积累量均为地上部>根系,即镉主要积累在黑麦草的地上部,分别占植株总镉积累量的64.1%~76.9%、64.6%~77.7%.比较2个黑麦草品种地上部、根系和植株总镉积累量,在各镉胁迫条件下均为’阿伯德‘>’邦德‘.
从表2可见,2个黑麦草品种地上部镉富集系数 (BCF) 随镉胁迫水平增加呈增长趋势,且镉富集系数均大于1,’邦德‘和’阿伯德‘在150~600 mg/kg Cd水平下的地上部BCF较75 mg/kg Cd水平下分别增加了162.8%~233.1%和25.3%~153.4%.2个黑麦草品种BCF根系>地上部(除’邦德‘150 mg/kg Cd水平);其中’阿伯德‘>’邦德‘(除600 mg/kg Cd水平外)。’邦德‘和’阿伯德‘转运系数在最大镉胁迫水平下达到最大值分别为3.33和3.49,且所有处理下TF值均大于1.黑麦草地上部和根系镉积累量以及植株总镉积累量在不同镉水平之间和不同黑麦草品种之间的差异均达到显着性水平 (P<0.05) .
表2 不同Cd水平对黑麦草各部位Cd积累量的影响
3 讨论与结论
镉是一种有毒的非必需元素,易于被植物吸收、富集,过量的镉会影响植物的正常生长发育[18].有研究表明,镉胁迫下植物的株高、生物量等生长指标会受到一定的抑制作用[19],对植物毒害的外观症状表现为株高矮小、生物产量下降、叶色减退、物候期延长、损伤植株根部、进而阻碍光合作用和蒸腾作用[20],且毒害作用随着Cd胁迫水平的增加而增加,严重时甚至会死亡。本试验中,在低Cd处理条件下,施加外源镉提高了’阿伯德‘地上部、根系干重和植株总干重,以及’邦德‘根系干重,这是因为在一定低Cd条件下,Cd可刺激和促进植物生长[21],这与韩航等[22]研究的低Cd处理对类芦分蘖数、叶片数、株高、最大叶长有一定促进作用,随Cd水平的增加,抑制作用逐渐增强等结果一致。比较2个品种黑麦草,在75~600 mg/kg Cd胁迫下,’阿伯德‘地上部干重和总干重均高于邦德品种,且’阿伯德‘品种在75 mg/kg Cd胁迫下,地上部干重高于对照,可见’阿伯德‘品种对镉有较强的的忍耐力,这与陈永勤[23]的研究结果一致。李慧芳等[24]研究指出根部对镉胁迫的敏感度明显高于叶片,低镉处理对地上部生物量会有促进作用,而根部会直线下降。随着镉处理水平的不断加大,其根系生物量降低的幅度也明显大于地上部分的变化幅度。而本试验中2个品种黑麦草根系干重呈先增加后降低的趋势,可见在较低水平Cd胁迫下,可以促进黑麦草根系生长,有利于其吸收营养和水分,进而提高植株的生物产量,表明黑麦草对Cd胁迫具有一定的抗性,这与的孙园园[25]的研究结果一致。
在镉胁迫下,植物体内活性氧 (Reactive oxygen species, ROS) 会大量增加,造成植物生理代谢障碍,影响植物正常生长。为了防止脂质过氧化损伤,植物通过提高SOD、POD、CAT等各种抗氧化酶的活性来维持植物体内ROS产生和清除的动态平衡[26,27,28,29].SOD酶可清除污染物诱导以及正常代谢过程产生的超氧阴离子自由基,是植物抗氧化系统发挥功能的首要酶[29],而CAT酶是过氧化氢酶体的主要酶类,POD酶是过氧化物酶体的主要酶类,均能将植物体内H2O2分解为O2和H2O.在本试验中,两个品种黑麦草地上部、根系SOD和CAT活性以及根系POD活性随着镉胁迫水平的增加呈先升高后降低的趋势,上升的原因是因为在低镉胁迫下,黑麦草耐受力较强,应激产生的保护作用以降低膜脂过氧化程度。随着镉水平的升高,镉引起的过氧化物不断积累,植株过氧化损伤不断加强,影响了植株正常的生理代谢,打破了植物体内ROS产生和清除的动态平衡,使抗氧化酶活性开始下降[30].2个品种黑麦草地上部POD活性随着镉胁迫水平的增加持续上升,可能是由于根系在高镉胁迫下清除体内H2O2的关键酶是POD,这与李冬琴等[31]研究结果一致。
通常逆境胁迫下植物的抗性与其体内MDA含量呈负相关[2].在本试验中,在镉胁迫下,’阿伯德‘地上部MDA含量先降后升,表明高镉胁迫下,过多的活性氧导致了膜脂过氧化,从而使得MDA含量上升[2,31].2个品种黑麦地上部MDA含量升高,其中’邦德‘品种MDA含量上升幅度大于’阿伯德‘品种,这说明’邦德‘品种在镉胁迫条件下的膜脂过氧化程度高于’阿伯德‘,可见’邦德‘抵抗不良环境的能力低于’阿伯德‘.根系MDA含量在高镉水平下呈下降趋势,可能是由于高水平镉胁迫对植株生长造成严重危害,也可能与植株对镉的耐受极限有关[32].
在一定浓度重金属污染土壤条件下,植物重金属富集系数、转移系数越大,表明其对重金属的吸收富集能力越强,越适合作为修复重金属污染土壤的植物材料。本试验中,2个品种黑麦草镉富集系数均大于1,且地上部镉富集系数在镉胁迫水平增加的情况下逐渐递增,根系镉富集系数也在最高镉胁迫水平下达到最大值,可见黑麦草各部位在高镉处理条件下积累能力较强,这与陈璐等[34]、冯鹏等[35]的研究结果一致。
本试验中,在75~600 mg/kg Cd胁迫范围内,黑麦草各部位Cd含量与Cd处理水平呈正相关关系;同时地上部和总植株镉积累量也逐渐升高,但是根系镉积累量在600 mg/kg Cd水平下反而下降,可能因为此镉胁迫下对根系生长抑制较强,根系生物量较小所致。比较’邦德‘和’阿伯德‘地上部和根系镉积累量可知,在相同Cd处理条件下,’邦德‘和’阿伯德‘地上部镉积累量均远大于根部,可见2个黑麦草品种地上部均比根系对重金属Cd有更强的蓄积能力。比较2个黑麦草品种还发现,’阿伯德‘地上部、根系镉含量基本高于’邦德‘,’阿伯德‘地上部、根系和植株总镉积累量也高于’邦德‘,可见’阿伯德‘对镉的富集能力高于邦德品种,同时也进一步印证了黑麦草对Cd耐性和吸收富集存在基因型差异,这与李慧芳等[24]和黄登峰等[13]的研究结果相似。
本试验中,随着土壤镉水平增加,2个黑麦草品种根系干重、’阿伯德‘地上部和植株总干重及地上部、根系SOD和CAT活性和根系POD活性呈先增加后降低的趋势。2个黑麦草品种地上部POD活性随着土壤镉水平的增加持续上升。随土壤镉水平增加,2个品种黑麦地上部MDA含量升高。’邦德‘和’阿伯德‘镉转运系数、镉富集系数均大于1.黑麦草对Cd耐性和吸收富集存在基因型差异,品种’阿伯德‘地上部干重和总干重均高于’邦德‘,且’阿伯德‘地上部和根系镉含量及镉积累量高于’邦德‘,’阿伯德‘更适合作为土壤镉污染的植物修复材料。
参考文献
[1] 熊仕娟。纳米沸石对Cd污染土壤的修复效应及机理研究[D].重庆:西南大学, 2016.
[2] 江玲。黑麦草、丛枝菌根对不同品种番茄镉吸收、富集的影响[D].重庆:西南大学, 2015.
[3] 郑莹辉, 熊仕娟, 徐卫红, 等。纳米沸石对大白菜镉吸收及土壤有效镉含量的影响[J].农业环境科学学报, 2016, 35 (12) :2353-2360.
[4] 秦余丽, 熊仕娟, 徐卫红, 等。纳米沸石对大白菜生长、抗氧化酶活性及镉形态、含量的影响[J].环境科学, 2017, 38 (3) :1189-1200.
[5] 周坤。外源锌、铁对番茄镉积累的影响研究[D].重庆:西南大学, 2014.
[6] Li J Y, Liu J, Dong D, et al.Natural variation underlies alterations in Nramp aluminum transporter (NRAT1) expression and function that play a key role in rice aluminum tolerance[J].Proceedings of the National Academy of Sciences of the United States of America, 2014, 111 (17) :6503-6508.
[7] 韦妮玉, 张新英。桂西北矿区农田土壤镉污染评价[J].绿色科技, 2016 (18) :31-33.
[8] 杨启良, 武振中, 陈金陵, 等。植物修复重金属污染土壤的研究现状及其水肥调控技术展望[J].生态环境学报, 2015 (6) :1075-1084.
[9] 李松克, 张春林, 李克勤, 等。多年生黑麦草对黄壤重金属污染的修复[J].贵州农业科学, 2014, 42 (11) :147-151.
[10] 孙园园, 关萍, 何杉, 等。镉胁迫对多花黑麦草镉积累特征、生理抗性及超微结构的影响[J].草业科学, 2016, 33 (8) :1589-1597.
[11] 李希铭。草本植物对镉的耐性和富集特征研究[D].北京:北京林业大学, 2016.
[12] 魏树强。多年生黑麦草 (Lolium perene L.) 耐镉机理与LpGCS基因的克隆和功能分析[D].北京:中国林业科学研究院, 2014.
[13] 黄登峰, 席嘉宾, 赵运林。镉胁迫下两个多年生黑麦草品种的生理响应[J].北方园艺, 2016 (3) :66-68.
[14] 唐让云, 曹靖, 董放, 等。淋洗与植物作用耦合对盐渍化土壤的改良效应[J].生态学报, 2015, 35 (17) :5686-5694.
[15] Liu Z, He Liu Z L, He X Y, et al.Accumulation and tolerance characteristics of cadmium in a potential hyperaccumulator:Lonicera japonica Thunb[J].Journal of Hazardous Materials, 2009, 169 (1) :170-175.
[16] 张志良。植物生理学实验指导 (第二版) [M].北京:高等教育出版社, 1990.
[17] 李合生。植物生理生化实验原理和技术[M].北京:高等教育出版社, 2000.
[18] 李志贤, 陈章, 陈国梁, 等。镉富集植物油菜与玉米间作对玉米吸收积累镉的影响[J].生态学杂志, 2016, 35 (1) :26-31.
[19] 吴福忠, 杨万勤, 张健, 等。镉胁迫对桂花生长和养分积累, 分配与利用的影响[J].植物生态学报, 2010, 34 (10) :1220-1226.
[20] 张兴梅, 杨清伟, 李扬。土壤镉污染现状及修复研究进展[J].河北农业科学, 2010, 03:79-81.
[21] 施宠, 王纯利, 黄长福, 等。镉胁迫对野燕麦幼苗生长及其生理特性的影响[J].草地学报, 2015, 23 (3) :526-532.
[22] 韩航, 陈雪娇, 侯晓龙, 等。镉胁迫对类芦生长及酶活性的影响[J].农业环境科学学报, 2016, 35 (4) :647-653.
[23] 陈永勤。镉富集植物镉积累基因型差异及分子机理研究[D].重庆:西南大学, 2017.
[24] 李慧芳, 王瑜, 袁庆华。镉胁迫对多年生黑麦草生长及生理特性的影响[J].中国草地学报, 2014, 36 (4) :79-84.
[25] 孙园园。耐镉植物抗性及富集规律的研究[D].贵阳:贵州大学, 2015.
[26] 王全辉, 梁雪, 董元杰, 等。外源NO对镉胁迫下黑麦草生长的缓解效应[J].中国草地学报, 2012, 34 (2) :30-35.
[27] Liu D L, Zhang S P, Chen Z, et al.Soil cadmium regulates antioxidases in sorghum[J].Agricultural Sciences in China, 2010, 9 (10) :1475-1480.
[28] 牛文婷。木炭对NaCl胁迫下小麦根系蛋白质组影响的研究[D].太原:山西师范大学, 2016.
[29] 孟利娟。一氧化氮和海藻糖对白灵侧耳高温响应抗氧化途径的影响研究[D].北京:中国农业科学院, 2015.
[30] 夏民旋, 王维, 袁瑞, 等。超氧化物歧化酶与植物抗逆性[J].分子植物育种, 2015, 13 (11) :2633-2646.
[31] 李冬琴, 陈桂葵, 郑海, 等。镉对两品种玉豆生长和抗氧化酶的影响[J].农业环境科学学报, 2015, 34 (2) :221-226.
[32] 张凯凯, 孙圆圆, 徐玲玲, 等。镉胁迫对一年生黑麦草生理生化特性的影响[J].山地农业生物学报, 2016, 35 (1) :86-89.
[33] 米艳华, 雷梅, 黎其万, 等。滇南矿区重金属污染耕地的植物修复及其健康风险[J].生态环境学报, 2016, 25 (5) :864-871
[34] 陈璐, 米艳华, 林昕, 等。土壤-三七系统重金属污染调查及相关分析[J].中国中药杂志, 2014, 39 (14) :2608.
[35] 冯鹏, 孙力, 申晓慧, 等。多年生黑麦草对Pb、Cd胁迫的响应及富集能力研究[J].草业学报, 2016, 25 (1) :153-162.